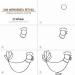
Искусственный нейрон
Схема искусственного нейрона
1.Нейроны, выходные сигналы которых поступают на вход данному
2.Сумматор входных сигналов
3.Вычислитель передаточной функции
4.Нейроны, на входы которых подаётся выходной сигнал данного
5. - веса
входных сигналов
Иску́сственный нейро́н (Математический нейрон Маккалока - Питтса , Формальный нейрон ) - узел искусственной нейронной сети , являющийся упрощённой моделью естественного нейрона . Математически, искусственный нейрон обычно представляют как некоторую нелинейную функцию от единственного аргумента - линейной комбинации всех входных сигналов. Данную функцию называют функцией активации или функцией срабатывания , передаточной функцией . Полученный результат посылается на единственный выход. Такие искусственные нейроны объединяют в сети - соединяют выходы одних нейронов с входами других. Искусственные нейроны и сети являются основными элементами идеального нейрокомпьютера .
Биологический прототип
Биологический нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро (с большим количеством ядерных пор) и другие органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами , аппарат Гольджи), и отростков. Выделяют два вида отростков. Аксон - обычно длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты - как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи с 20-ю тысячами других нейронов. Кора головного мозга человека содержит 10-20 миллиардов нейронов.
История развития
При этом возможен сдвиг функции по обеим осям (как изображено на рисунке).
Недостатками шаговой и полулинейной активационных функций относительно линейной можно назвать то, что они не являются дифференцируемыми на всей числовой оси, а значит не могут быть использованы при обучении по некоторым алгоритмам.

Пороговая функция активации
Пороговая передаточная функция
Гиперболический тангенс
Здесь - расстояние между центром и вектором входных сигналов . Скалярный параметр определяет скорость спадания функции при удалении вектора от центра и называется шириной окна , параметр определяет сдвиг активационной функции по оси абсцисс. Сети с нейронами, использующими такие функции, называются RBF-сетями. В качестве расстояния между векторами могут быть использованы различные метрики , обычно используется евклидово расстояние:
Здесь - j-я компонента вектора, поданного на вход нейрона, а - j-я компонента вектора, определяющего положение центра передаточной функции. Соответственно, сети с такими нейронами называются вероятностными и регрессионными .
В реальных сетях активационная функция этих нейронов может отражать распределение вероятности какой-либо случайной величины , либо обозначать какие-либо эвристические зависимости между величинами.
Другие функции передачи
Перечисленные выше функции составляют лишь часть от множества передаточных функций, используемых на данный момент. В число других передаточных функций входят такие как :
Стохастический нейрон
Выше описана модель детерминистического искусственного нейрона, то есть состояние на выходе нейрона однозначно определено результатом работы сумматора входных сигналов. Рассматривают также стохастические нейроны, где переключение нейрона происходит с вероятностью, зависящей от индуцированного локального поля, то есть передаточная функция определена как
где распределение вероятности обычно имеет вид сигмоида
a нормировочная константа вводится для условия нормализации распределения вероятности . Таким образом, нейрон активируется с вероятностью P(u). Параметр T - аналог температуры (но не температуры нейрона!) и определяет беспорядок в нейронной сети. Если Т устремить к 0, стохастический нейрон перейдет в обычный нейрон с передаточной функцией Хевисайда (пороговой функцией).
Моделирование формальных логических функций
Нейрон с пороговой передаточной функцией может моделировать различные логические функции. Изображения иллюстрируют, каким образом можно, задав веса входных сигналов и порог чувствительности, заставить нейрон выполнять конъюнкцию (логическое «И») и дизъюнкцию (логическое «ИЛИ») над входными сигналами, а также логическое отрицание входного сигнала . Этих трех операций достаточно, чтобы смоделировать абсолютно любую логическую функцию любого числа аргументов.
Примечания
Литература
- Терехов В.А., Ефимов Д.В., Тюкин И.Ю. Нейросетевые системы управления. - 1-е. - Высшая школа, 2002. - С. 184. - ISBN 5-06-004094-1
- Круглов В. В., Борисов В. В.
Рис.
1.2.
На рис. 1.2 представлена модель, реализующая эту идею. Множество входных сигналов, обозначенных , поступает на искусственный нейрон . Эти входные сигналы, в совокупности обозначаемые вектором , соответствуют сигналам, приходящим в синапсы биологического нейрона . Каждый сигнал умножается на соответствующий вес , и поступает на суммирующий блок, обозначенный Каждый вес соответствует "силе" одной биологической синаптической связи. (Множество весов в совокупности обозначается вектором ) Суммирующий блок, соответствующий телу биологического элемента, складывает взвешенные входы алгебраически, создавая выход , который мы будем называть В векторных обозначениях это может быть компактно записано следующим образом:
где - константа, пороговой функцией

где - некоторая постоянная пороговая величина, или же функция , более точно моделирующая нелинейную передаточную характеристику биологического нейрона и предоставляющей нейронной сети большие возможности.

Рис. 1.3.
На рис. 1.2 блок, обозначенный , принимает сигнал и выдает сигнал Если блок сужает диапазон изменения величины так, что при любых значениях значения принадлежат некоторому конечному интервалу, то называется "сжимающей" функцией . В качестве "сжимающей" функции часто используется логистическая или "сигмоидальная" (S-образная) функция , показанная на рис. 1.3 . Эта функция математически выражается как Таким образом,

По аналогии с электронными системами активационную функцию можно считать нелинейной усилительной характеристикой искусственного нейрона . Коэффициент усиления вычисляется как отношение приращения величины к вызвавшему его небольшому приращению величины Он выражается наклоном кривой при определенном уровне возбуждения и изменяется от малых значений при больших отрицательных возбуждениях (кривая почти горизонтальна) до максимального значения при нулевом возбуждении и снова уменьшается, когда возбуждение становится большим положительным. С. Гроссберг (1973) обнаружил, что подобная нелинейная характеристика решает поставленную им дилемму шумового насыщения. Каким образом одна и та же сеть может обрабатывать как слабые, так и сильные сигналы? Слабые сигналы нуждаются в большом сетевом усилении, чтобы дать пригодный к использованию выходной сигнал. Однако усилительные каскады с большими коэффициентами усиления могут привести к насыщению выхода шумами усилителей (случайными флуктуациями), которые присутствуют в любой физически реализованной сети. Сильные входные сигналы, в свою очередь , также будут приводить к насыщению усилительных каскадов, исключая возможность полезного использования выхода. Центральная область логистической функции, имеющая большой коэффициент усиления, решает проблему обработки слабых сигналов, в то время как области с падающим усилением на положительном и отрицательном концах подходят для больших возбуждений. Таким образом, нейрон функционирует с большим усилением в широком диапазоне уровня входного сигнала
Другой широко используемой
Адаптивные информационные системы
Потребность в адаптивных информационных системах возникает в тех случаях, когда поддерживаемые ими проблемные области постоянно развиваются. В связи с этим адаптивные системы должны удовлетворять ряду специфических требований, а именно:
Адекватно отражать знания проблемной области в каждый момент времени;
Быть пригодными для легкой и быстрой реконструкции при изменении проблемной среды.
Адаптивные свойства информационных систем обеспечиваются за счет интеллектуализации их архитектуры. Ядром таких систем является постоянно развиваемая модель проблемной области, поддерживаемая в специальной базе знаний – репозитории. Ядро системы управляет процессами генерации или переконфигурирования программного обеспечения.
В процессе разработки адаптивных информационных систем применяется оригинальное или типовое проектирование. Оригинальное проектирование предполагает разработку информационной системы с «чистого листа» на основе сформулированных требований. Реализация этого подхода основана на использовании систем автоматизированного проектирования, или CASE-технологий (Designer2000, SilverRun, Natural Light Storm и др.).
При типовом проектировании осуществляется адаптация типовых разработок к особенностям проблемной области. Для реализации этого подхода применяются инструментальные средства компонентного (сборочного) проектирования информационных систем (R/3, BAAN IV, Prodis и др.).
Главное отличие подходов состоит в том, что при использовании CASE-технологии каждый раз при изменении проблемной области выполняется генерация программного обеспечения в целом, а при использовании сборочной технологии – конфигурирование модулей и только в редких случаях – их переработка.
Тема 2. Искусственные нейронные сети. Архитектура и классификация нейронных сетей.
Представление о детальном устройстве головного мозга появилось только около ста лет назад. В 1888 г. испанский доктор Рамони Кайал экспериментально показал, что мозговая ткань состоит из большого числа связанных друг с другом однотипных узлов – нейронов. Более поздние исследования при помощи электронного микроскопа показали, что все нейроны, независимо от типа, имеют схожую организационную структуру (рис. 2.1). Естественная нервная клетка (нейрон) состоит из тела (сомы), содержащего ядро, и отростков – дендритов, по которым в нейрон поступают входные сигналы. Один из отростков, ветвящийся на конце, служит для передачи выходных сигналов данного нейрона другим нервным клеткам. Он называется аксоном. Соединение аксона с дендритом другого нейрона называется синапсом. Нейрон возбуждается и передает сигнал через аксон, если число пришедших по дендритам возбуждающих сигналов больше, чем число тормозящих.
Рисунок 2.1 – Строение биологического нейрона.
В 1943 г. В. Маккаллох и В. Питтс предложили систему обработки информации в виде сети, состоящей из простых вычислителей, созданных по принципу биологического нейрона. Искусственная нейронная сеть (ИНС) представляет собой совокупность простых вычислительных элементов (процессоров) – искусственных нейронов, соединенных некоторым образом так, чтобы между ними обеспечивалось взаимодействие. Искусственные нейроны характеризуются правилом комбинирования входных сигналов и передаточной функцией, позволяющим вычислить выходной сигнал.

Рисунок 2.2 – Кибернетическая модель нейрона.
Информация, поступающая на вход нейрона, суммируется с учетом весовых коэффициентов сигналов:
![]() , (2.1)
, (2.1)
где w 0 – сдвиг (порог, смещение) нейрона.
В зависимости от значения весового коэффициента w i , входной сигнал x i или усиливается, или подавляется. Взвешенную сумму входных сигналов также называют потенциалом или комбинированным входом нейрона.
Сдвиг обычно интерпретируется как связь, исходящая от элемента, активность которого всегда равна 1. Обычно, для удобства входной вектор расширяется добавлением этого сигнала до х = (1,х 0 ,...,x n) и порог w 0 вносится под знак суммы:
Передаточная функция, или функция активации нейрона, является правилом, согласно которому взвешенная сумма поступивших сигналов P преобразуется в выходной сигнал нейрона Y, который передается другим нейронам сети, т. е. Y=f(P). На рисунке 2.3 показаны графики наиболее распространённых функций активации нейронов.
Пороговая функция пропускает информацию только в том случае, если алгебраическая сумма входных сигналов превышает некоторую постоянную величину Р*, например:

Пороговая функция не обеспечивает достаточной гибкости ИНС при обучении. Если значение вычисленного потенциала не достигает заданного порога, то выходной сигнал не формируется и нейрон «не срабатывает». Это приводит к снижению интенсивности выходного сигнала нейрона и, как следствие, к формированию невысокого значения потенциала взвешенных входов в следующем слое нейронов.
Линейная функция дифференцируема и легко вычисляется, что в ряде случаев позволяет уменьшить ошибки выходных сигналов в сети, так как передаточная функция сети также является линейной. Однако она не универсальна и не обеспечивает решения многих задач.
Определенным компромиссом между линейной и ступенчатой функциями является сигмоидальная функция активации Y = 1/(1+exp(-kP)), которая удачно моделирует передаточную характеристику биологического нейрона (рис. 3.3, в).

Рисунок 2.3 – Функции переноса искусственных нейронов:
а) линейная; б) ступенчатая; в) сигмоидальная.
Коэффициент k определяет крутизну нелинейной функции: чем больше k, тем ближе сигмоидальная функция к пороговой; чем меньше k, тем она ближе k линейной. Тип функции переноса выбирается с учетом конкретной задачи, решаемой с применением нейронных сетей. Например, в задачах аппроксимации и классификации предпочтение отдают сигмоидальной кривой.
Архитектура и классификация ИНС
С каждым нейроном связывается набор входящих связей, по которым к данному элементу поступают сигналы от других элементов сети, и набор исходящих связей, по которым сигналы данного элемента передаются другим нейронам. Некоторые нейроны предназначены для получения сигналов из внешней среды (входные элементы), а некоторые – для вывода во внешнюю среду результатов вычислений (выходные элементы).
В 1958 г. Фрэнк Розенблатт предложил следующую модель нейронной сети – персептрона. Персептрон Розенблатта (рис. 2.4) состоит из k нейронов, имеет d входов, k выходов и только один слой настраиваемых весов w ij .

Рисунок 2.4 – Персептрон Розенблатта.
Входные нейроны обычно предназначены для распределения вводимых сигналов между другими нейронами сети, поэтому для них требуется, чтобы исходящий от элемента сигнал был таким же, как и входящий. В отличие от других нейронов сети, входные имеют только по одному входу. Иными словами, каждый входной элемент может получать сигнал от одного соответствующего ему датчика. Поскольку входные элементы предназначены исключительно для того, чтобы распределять сигналы, получаемые из внешней среды, многие исследователи вообще не считают входные элементы частью нейронной сети.
Персептрон способен решать линейные задачи. Число входов сети определяет размерность пространства, из которого выбираются входные данные: для двух признаков пространство оказывается двумерным, для трех – трехмерным, а для d признаков – d-мерным. Если прямая или гиперплоскость в пространстве входных данных может разделить все образцы на соответствующие им классы, то проблема является линейной, в противном случае – нелинейной. На рисунке 2.5 показаны множества точек на плоскости, причём в случае а) граница линейная, в случае – б) нелинейная.

Рисунок 2.5 – Геометрическое представление линейной (а) и
нелинейной (б) задач.
Для решения нелинейных проблем предложены модели многослойных персептронов (MLP), способные строить ломаную границу между распознаваемыми образами. В многослойных сетях каждый нейрон может посылать выходной сигнал только в следующий слой и принимать входные сигналы только с предыдущего слоя, как показано на рисунке 2.6. Слои нейронов, расположенные между входным и выходным называются скрытыми, так как не получают и не передают данные непосредственно из внешней среды. Такая сеть позволяет выделять глобальные свойства данных за счет наличия дополнительных синаптических связей и повышения уровня взаимодействия нейронов.

Рисунок 2.6 – Схема многослойного персептрона.
Определение числа скрытых слоев и числа нейронов в каждом слое для конкретной задачи является неформальной проблемой, при решении которой можно использовать эвристическое правило: число нейронов в следующем слое в два раза меньше, чем в предыдущем
В настоящее время кроме многослойного персептрона существует множество способов задания структур нейронных сетей. Все виды нейронных сетей можно условно разделить на сети прямого распространения и сети с обратными связями. Как следует из названия, в сетях первого типа сигналы от нейрона к нейрону распространяются в четко заданном направлении – от входов сети к ее выходам. В сетях второго типа выходные значения любого нейрона сети могут передаваться к его же входам. Это позволяет нейронной сети моделировать более сложные процессы, например временные, но делает выходы подобной сети нестабильными, зависящими от состояния сети на предыдущем цикле. На рисунке 2.7. представлена классификация наиболее распространенных типов нейронных сетей.

Рисунок 2.7 – Классификация распространённых видов ИНС.
Тема 3. Сети типа персептрон
Задача классификации образов. Обучение с учителем
Ф. Розенблатт предложил использовать персептрон для задач классификации. Многие приложения можно интерпретировать, как проблемы классификации. Например, оптическое распознавании символов. Отсканированные символы ассоциируются с соответствующими им классами. Имеется немало вариантов изображения буквы "Н" даже для одного конкретного шрифта – символ может оказаться, например, смазанным, – но все эти изображения должны принадлежать классу "Н".
Когда известно, к какому классу относится каждый из учебных примеров, можно использовать стратегию обучения с учителем. Задачей для сети является ее обучение тому, как сопоставить предъявляемый сети образец с контрольным целевым образцом, представляющим нужный класс. Иными словами, знания об окружающей среде представляются нейронной сети в виде пар "вход-выход". Например, сети можно предъявить изображение буквы "Н" и обучить сеть тому, что при этом соответствующий "Н" выходной элемент должен быть включен, а выходные элементы, соответствующие другим буквам – выключены. В этом случае входной образец может быть набором значений, характеризующих пиксели изображения в оттенках серого, а целевой выходной образец – вектором, значения всех координат которого должны быть равными 0, за исключением координаты, соответствующей классу "Н", значение которой должно быть равным.
На рисунке 3.1 показана блочная диаграмма, иллюстрирующая эту форму обучения. Предположим, что учителю и обучаемой сети подается обучающий вектор из окружающей среды. На основе встроенных знаний учитель может сформировать и передать обучаемой нейронной сети желаемый отклик, соответствующий данному входному вектору. Параметры сети корректируются с учетом обучающего вектора и сигнала ошибки. Сигнал ошибки – это разность между желаемым сигналом и текущим откликом нейронной сети. После окончания обучения учителя можно отключить и позволить нейронной сети работать со средой самостоятельно.

Рисунок 3.1 – Концепция обучения ИНС с учителем.
Алгоритм обучения перцептрона включает следующие шаги:
· Системе предъявляется эталонный образ.
· Если результат распознавания совпадает с заданным, весовые коэффициенты связей не изменяются.
· Если ИНС неправильно распознает результат, то весовым коэффициентам дается приращение в сторону повышения качества распознавания.
Была представлена модель нервной системы, опишу теорию и принципы, которые легли в её основу.
Теория основана на анализе имеющейся информации о биологическом нейроне и нервной системе из современной нейробиологии и физиологии мозга.
Сначала приведу краткую информацию об объекте моделирования, вся информация изложена далее, учтена и использована в модели.
НЕЙРОН
Нейрон является основным функциональным элементом нервной системы, он состоит из тела нервной клетки и её отростков. Существуют два вида отростков: аксоны и дендриты. Аксон – длинный покрытый миелиновой оболочкой отросток, предназначенный для передачи нервного импульса на далекие расстояния. Дендрит – короткий, ветвящийся отросток, благодаря которым происходит взаимосвязь с множеством соседних клеток.
ТРИ ТИПА НЕЙРОНОВ
Нейроны могут сильно отличаться по форме, размерам и конфигурации, не смотря на это, отмечается принципиальное сходство нервной ткани в различных участках нервной системе, отсутствуют и серьезные эволюционные различия. Нервная клетка моллюска Аплизии может выделять такие же нейромедиаторы и белки, что и клетка человека.В зависимости от конфигурации выделяют три типа нейронов:
А) рецепторные, центростремительные, или афферентные нейроны, данные нейроны имеют центростремительный аксон, на конце которого имеются рецепторы, рецепторные или афферентные окончания. Эти нейроны можно определить, как элементы, передающие внешние сигналы в систему.
Б) интернейроны (вставочные, контактные, или промежуточные) нейроны, не имеющие длинных отростков, но имеющие только дендриты. Таких нейронов в человеческом мозгу больше чем остальных. Данный вид нейронов является основным элементом рефлекторной дуги.
В) моторные, центробежные, или эфферентные, они имеют центростремительный аксон, который имеет эфферентные окончания передающий возбуждение мышечным или железистым клеткам. Эфферентные нейроны служат для передачи сигналов из нервной среды во внешнюю среду.
Обычно в статьях по искусственным нейронным сетям оговаривается наличие только моторных нейронов (с центробежным аксоном), которые связаны в слои иерархической структуры. Подобное описание применимо к биологической нервной системе, но является своего рода частным случаем, речь идет о структурах, базовых условных рефлексов. Чем выше в эволюционном значении нервная система, тем меньше в ней превалируют структуры типа «слои» или строгая иерархия.
ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ
Передача возбуждения происходит от нейрона к нейрону, через специальные утолщения на концах дендритов, называемых синапсами. По типу передачи синапсы разделяют на два вида: химические и электрические. Электрические синапсы передают нервный импульс непосредственно через место контакта. Таких синапсов в нервных системах очень мало, в моделях не будут учитываться. Химические синапсы передают нервный импульс посредством специального вещества медиатора (нейромедиатора, нейротрансмиттера), данный вид синапса широко распространен и подразумевает вариативность в работе.Важно отметить, что в биологическом нейроне постоянно происходят изменения, отращиваются новые дендриты и синапсы, возможны миграции нейронов. В местах контактов с другими нейронами образуются новообразования, для передающего нейрона - это синапс, для принимающего - это постсинаптическая мембрана, снабжаемая специальными рецепторами, реагирующими на медиатор, то есть можно говорить, что мембрана нейрона - это приемник, а синапсы на дендритах - это передатчики сигнала.
СИНАПС

При активации синапса он выбрасывает порции медиатора, эти порции могут варьироваться, чем больше выделится медиатора, тем вероятнее, что принимаемая сигнал нервная клетка будет активирована. Медиатор, преодолевая синоптическую щель, попадает на постсинаптическую мембрану, на которой расположены рецепторы, реагирующие на медиатор. Далее медиатор может быть разрушен специальным разрушающим ферментом, либо поглощен обратно синапсом, это происходит для сокращения времени действия медиатора на рецепторы.
Так же помимо побудительного воздействия существуют синапсы, оказывающие тормозящее воздействие на нейрон. Обычно такие синапсы принадлежат определенным нейронам, которые обозначаются, как тормозящие нейроны.
Синапсов связывающих нейрон с одной и той же целевой клеткой, может быть множество. Для упрощения примем, всю совокупность, оказываемого воздействия одним нейроном, на другой целевой нейрон за синапс с определённой силой воздействия. Главной характеристикой синапса будет, является его сила.
СОСТОЯНИЕ ВОЗБУЖДЕНИЯ НЕЙРОНА
В состоянии покоя мембрана нейрона поляризована. Это означает, что по обе стороны мембраны располагаются частицы, несущие противоположные заряды. В состоянии покоя наружная поверхность мембраны заряжена положительно, внутренняя – отрицательно. Основными переносчиками зарядов в организме являются ионы натрия (Na+), калия (K+) и хлора (Cl-).Разница между зарядами поверхности мембраны и внутри тела клетки составляет мембранный потенциал. Медиатор вызывает нарушения поляризации – деполяризацию. Положительные ионы снаружи мембраны устремляются через открытые каналы в тело клетки, меняя соотношение зарядов между поверхностью мембраны и телом клетки.

Изменение мембранного потенциала при возбуждении нейрона
Характер изменений мембранного потенциала при активации нервной ткани неизменен. Независимо от того кокой силы воздействия оказывается на нейрон, если сила превышает некоторое пороговое значение, ответ будет одинаков.
Забегая вперед, хочу отметить, что в работе нервной системы имеет значение даже следовые потенциалы (см. график выше). Они не появляются, вследствие каких-то гармонических колебаний уравновешивающих заряды, являются строгим проявлением определённой фазы состояния нервной ткани при возбуждении.
ТЕОРИЯ ЭЛЕКТРОМАГНИТНОГО ВЗАИМОДЕЙСТВИЯ
Итак, далее приведу теоретические предположения, которые позволят нам создавать математические модели. Главная идея заключается во взаимодействии между зарядами формирующихся внутри тела клетки, во время её активности, и зарядами с поверхностей мембран других активных клеток. Данные заряды являются разноименными, в связи этим можно предположить, как будут располагаться заряды в теле клетки под воздействием зарядов других активных клеток.
Можно сказать, что нейрон чувствует активность других нейронов на расстоянии, стремится направить распространения возбуждения в направлении других активных участков.
В момент активности нейрона можно рассчитать определённую точку в пространстве, которая определялась бы, как сумма масс зарядов, расположенных на поверхностях других нейронов. Указанную точку назовем точкой паттерна, её месторождение зависит от комбинации фаз активности всех нейронов нервной системы. Паттерном в физиологии нервной системы называется уникальная комбинация активных клеток, то есть можно говорить о влиянии возбуждённых участков мозга на работу отдельного нейрона.
Нужно представлять работу нейрона не просто как вычислителя, а своего рода ретранслятор возбуждения, который выбирает направления распространения возбуждения, таким образом, формируются сложные электрические схемы. Первоначально предполагалось, что нейрон просто избирательно отключает/включает для передачи свои синапсы, в зависимости от предпочитаемого направления возбуждения. Но более детальное изучение природы нейрона, привело к выводам, что нейрон может изменять степень воздействия на целевую клетку через силу своих синапсов, что делает нейрон более гибким и вариативным вычислительным элементом нервной системы.

Какое же направление для передачи возбуждения является предпочтительным? В различных экспериментах связанных с образованием безусловных рефлексов, можно определить, что в нервной системе образуются пути или рефлекторные дуги, которые связывают активируемые участки мозга при формировании безусловных рефлексов, создаются ассоциативные связи. Значит, нейрон должен передавать возбуждения к другим активным участкам мозга, запоминать направление и использовать его в дальнейшем.
Представим вектор начало, которого находится в центре активной клети, а конец направлен в точку паттерна определённую для данного нейрона. Обозначим, как вектор предпочитаемого направления распространения возбуждения (T, trend). В биологическом нейроне вектор Т может проявляться в структуре самой нейроплазмы, возможно, это каналы для движения ионов в теле клетки, или другие изменения в структуре нейрона.
Нейрон обладает свойством памяти, он может запоминать вектор Т, направление этого вектора, может меняться и перезаписываться в зависимости от внешних факторов. Степень с которой вектор Т может подвергается изменениям, называется нейропластичность.
Этот вектор в свою очередь оказывает влияние на работу синапсов нейрона. Для каждого синапса определим вектор S начало, которого находится в центре клетки, а конец направлен в центр целевого нейрона, с которым связан синапс. Теперь степень влияния для каждого синапса можно определить следующим образом: чем меньше угол между вектором T и S, тем больше синапс будет, усиливается; чем меньше угол, тем сильнее синапс будет ослабевать и возможно может прекратить передачу возбуждения. Каждый синапс имеет независимое свойство памяти, он помнит значение своей силы. Указанные значения изменяются при каждой активизации нейрона, под влиянием вектора Т, они либо увеличиваются, либо уменьшаются на определённое значение.
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ

Входные сигналы (x1, x2,…xn) нейрона представляют собой вещественные числа, которые характеризуют силу синапсов нейронов, оказывающих воздействие на нейрон.
Положительное значение входа означает побудительное воздействие, оказываемое на нейрон, а отрицательное значение – тормозящее воздействие.
Для биологического нейрона не имеет значение, откуда поступил возбуждающий его сигнал, результат его активности будет идентичен. Нейрон будет активизирован, когда сумма воздействий на него будет превышать определённое пороговое значение. Поэтому, все сигналы проходят через сумматор (а), а поскольку нейроны и нервная система работают в реальном времени, следовательно, воздействие входов должно оцениваться в короткий промежуток времени, то есть воздействие синапса имеет временный характер.
Результат сумматора проходит пороговую функцию (б), если сумма превосходит пороговое значение, то это приводит к активности нейрона.
При активации нейрон сигнализирует о своей активности системе, передовая информацию о своём положении в пространстве нервной системы и заряде, изменяемом во времени (в).
Через определённое время, после активации нейрон передает возбуждение по всем имеющимся синапсам, предварительно производя пересчет их силы. Весь период активации нейрон перестает реагировать на внешние раздражители, то есть все воздействия синапсов других нейронов игнорируются. В период активации входит так же период восстановления нейрона.
Происходит корректировка вектора Т (г) с учётом значения точки паттерна Pp и уровнем нейропластичности. Далее происходит переоценка значений всех сил синапсов в нейроне(д).
Обратите внимание, что блоки (г) и (д) выполняются параллельно с блоком (в).
ЭФФЕКТ ВОЛНЫ
Если внимательно проанализировать предложенную модель, то можно увидеть, что источник возбуждения должен оказывать большее влияние на нейрон, чем другой удалённый, активный участок мозга. Следовательно возникает вопрос: почему же все равно происходит передача в направлении другого активного участка?Данную проблему я смог определить, только создав компьютерную модель. Решение подсказал график изменения мембранного потенциала при активности нейрона.

Усиленная реполяризация нейрона, как говорилось ранее, имеет важное значение для нервной системы, благодаря ей создается эффект волны, стремление нервного возбуждения распространятся от источника возбуждения.
При работе с моделью я наблюдал два эффекта, ели пренебречь следовым потенциалом или сделать его недостаточно большим, то возбуждение не распространяется от источников, а в большей степени стремится к локализации. Если сделать следовой потенциал сильно большим, то возбуждение стремится «разбежаться» в разные стороны, не только от своего источника, но и от других.
КОГНИТИВНАЯ КАРТА
Используя теорию электромагнитного взаимодействия, можно объяснить многие явления и сложные процессы, протекающие в нервной системе. К примеру, одним из последних открытий, которое широко обсуждается в науках о мозге, является открытие когнитивных карт в гиппокампе.Гиппокамп – это отдел мозга, которому отвечает за кратковременную память. Эксперименты на крысах выявили, что определённому месту в лабиринте соответствует своя локализованная группа клеток в гиппокампе, причем, не имеет значение, как животное попадает в это место, все равно будет активирован соответствующий этому месту участок нервной ткани. Естественно, животное должно помнить данный лабиринт, не стоит рассчитывать на топологическое соответствие пространства лабиринта и когнитивной карты.

Каждое место в лабиринте представляется в мозге, как совокупность раздражителей различного характера: запахи, цвет стен, возможные примечательные объекты, характерные звуки и т. д. Указанные раздражители отражаются на коре, различных представительствах органов чувств, в виде всплесков активности в определённых комбинациях. Мозг одновременно обрабатывает информацию в нескольких отделах, зачастую информационные каналы разделяются, одна и та же информация поступает в различные участки мозга.

Активация нейронов места в зависимости от положения в лабиринте (активность разных нейронов показана разным цветом).
Гиппокамп расположен в центре мозга, вся кара и её области удалены от него, на одинаковые расстояния. Если определить для каждой уникальной комбинации раздражителей точку масс зарядов поверхностей нейронов, то можно увидеть, что указанные точки будут различны, и будут находиться примерно в центре мозга. К этим точкам будет стремиться и распространятся возбуждение в гиппокампе, формируя устойчивые участки возбуждения. Более того, поочередная смена комбинаций раздражителей, будет приводить к смещению точки паттерна. Участки когнитивной карты будут ассоциативно связываться друг с другом последовательно, что приведет к тому, что животное, помещенное в начало знакомого ей лабиринта, может вспомнить весь последующий путь.
Заключение
У многих возникнет вопрос, где в данной работе предпосылки к элементу разумности или проявления высшей интеллектуальной деятельности?Важно отметить, что феномен человеческого поведения, есть следствие функционирования биологической структуры. Следовательно, чтобы имитировать разумное поведение, необходимо хорошо понимать принципы и особенности функционирования биологических структур. К сожалению, в науке биологии пока не представлен четкий алгоритм: как работает нейрон, как понимает, куда необходимо отращивать свои дендриты, как настроить свои синапсы, что бы в нервной системе смог сформироваться простой условный рефлекс, на подобие тех, которые демонстрировал и описывал в своих работах академик И.П. Павлов.
С другой стороны в науке об искусственном интеллекте, в восходящем (биологическом) подходе, сложилось парадоксальная ситуация, а именно: когда используемые в исследованиях модели основаны на устаревших представлениях о биологическом нейроне, консерватизм, в основе которого берётся персептрон без переосмысления его основных принципов, без обращения к биологическому первоисточнику, придумывается все более хитроумные алгоритмы и структуры, не имеющих биологических корней.
Конечно, никто не уменьшает достоинств классических нейронных сетей, которые дали множество полезных программных продуктов, но игра с ними не является путем к созданию интеллектуально действующей системы.
Более того, не редки заявления, о том, что нейрон подобен мощной вычислительной машине, приписывают свойство квантовых компьютеров. Из-за этой сверхсложности, нервной системе приписывается невозможность её повторения, ведь это соизмеримо с желанием смоделировать человеческую душу. Однако, в реальности природа идет по пути простоты и элегантности своих решений, перемещение зарядов на мембране клетки может служить, как для передачи нервного возбуждения, так и для трансляции информации о том, где происходит данная передача.
Несмотря на то, что указанная работа демонстрирует, как образуются элементарные условные рефлексы в нервной системе, она приближает к пониманию того, что такое интеллект и разумная деятельность.
Существуют еще множество аспектов работы нервной системы: механизмы торможения, принципы построения эмоций, организация безусловных рефлексов и обучение, без которых невозможно построить качественную модель нервной системы. Есть понимание, на интуитивном уровне, как работает нервная система, принципы которой возможно воплотить в моделях.
Создание первой модели помогли отработать и откорректировать представление об электромагнитном взаимодействии нейронов. Понять, как происходит формирование рефлекторных дуг, как каждый отдельный нейрон понимает, каким образом ему настроить свои синапсы для получения ассоциативных связей.
На данный момент я начал разрабатывать новую версию программы, которая позволит смоделировать многие другие аспекты работы нейрона и нервной системы.
Прошу принять активное участие в обсуждении выдвинутых здесь гипотез и предположений, так как я могу относиться к своим идеям предвзято. Ваше мнение очень важно для меня.
Теги:
- нейронные сети
- искусственный интеллект
- мозг
Искусственный нейрон имитирует в первом приближении свойства биологического нейрона. На вход искусственного нейрона поступает некоторое множество сигналов, каждый из которых является выходом другого нейрона. Каждый вход умножается на соответствующий вес, аналогичный синаптической силе, и все произведения суммируются, определяя уровень активации нейрона. На рис. 1.2 представлена модель, реализующая эту идею. Хотя сетевые парадигмы весьма разнообразны, в основе почти всех их лежит эта конфигурация. Здесь множество входных сигналов, обозначенных x 1 ,x 2 , …, x n , поступает на искусственный нейрон. Эти входные сигналы, в совокупности обозначаемые векторомX , соответствуют сигналам, приходящим в синапсы биологического нейрона. Каждый сигнал умножается на соответствующий вес w 1 , w 2 , …, w n , и поступает на суммирующий блок, обозначенный Σ.Каждый вес соответствует «силе» одной биологической синаптической связи. (Множество весов в совокупности обозначается векторомW .) Суммирующий блок, соответствующий телу биологического элемента, складывает взвешенные входы алгебраически, создавая выход, который мы будем называть NET.В векторных обозначениях это может быть компактно записано следующим образом:
NET =XW .
Рис. 1.2. Искусственный нейрон
Активационные функции
OUT= K (NET),
где К – постоянная, пороговой функции
OUT = 1, если NET > T, OUT = 0 в остальных случаях,
где Т – некоторая постоянная пороговая величина, или же функцией, более точно моделирующей нелинейную передаточную характеристику биологического нейрона и представляющей нейронной сети большие возможности.

Рис. 1.3. Искусственный нейрон с активационной функцией
На рис. 1.3 блок, обозначенный F, принимает сигналNETи выдает сигнал OUT.Если блокF сужает диапазон изменения величины NETтак, что при любых значениях NET значения OUTпринадлежат некоторому конечному интервалу, тоF называется«сжимающей» функцией. В качестве «сжимающей» функции часто используется логистическая или «сигмоидальная» (S-образная) функция, показанная на рис. 1.4а. Эта функция математически выражается какF(x) = 1/(1 + е - x). Таким образом,
 .
.
По аналогии с электронными системами активационную функцию можно считать нелинейной усилительной характеристикой искусственного нейрона. Коэффициент усиления вычисляется как отношение приращения величины OUTк вызвавшему его небольшому приращению величины NET.Он выражается наклоном кривой при определенном уровне возбуждения и изменяется от малых значений при больших отрицательных возбуждениях (кривая почти горизонтальна) до максимального значения при нулевом возбуждении и снова уменьшается, когда возбуждение становится большим положительным. Гроссберг (1973) обнаружил, что подобная нелинейная характеристика решает поставленную им дилемму шумового насыщения. Каким образом одна и та же сеть может обрабатывать как слабые, так и сильные сигналы? Слабые сигналы нуждаются в большом сетевом усилении, чтобы дать пригодный к использованию выходной сигнал. Однако усилительные каскады с большими коэффициентами усиления могут привести к насыщению выхода шумами усилителей (случайными флуктуациями), которые присутствуют в любой физически реализованной сети. Сильные входные сигналы в свою очередь также будут приводить к насыщению усилительных каскадов, исключая возможность полезного использования выхода. Центральная область логистической функции, имеющая большой коэффициент усиления, решает проблему обработки слабых сигналов, в то время как области с падающим усилением на положительном и отрицательном концах подходят для больших возбуждений. Таким образом, нейрон функционирует с большим усилением в широком диапазоне уровня входного сигнала.
 .
.

Рис. 1.4а. Сигмоидальная логистическая функция
Другой широко используемой активационной функцией является гиперболический тангенс. По форме она сходна с логистической функцией и часто используется биологами в качестве математической модели активации нервной клетки. В качестве активационной функции искусственной нейронной сети она записывается следующим образом:

Рис. 1.4б. Функция гиперболического тангенса
Подобно логистической функции гиперболический тангенс является S-образной функцией, но он симметричен относительно начала координат, и в точке NET= 0 значение выходного сигнала OUTравно нулю (см. рис. 1.4б). В отличие от логистической функции гиперболический тангенс принимает значения различных знаков, что оказывается выгодным для ряда сетей (см. гл. 3).
Рассмотренная простая модель искусственного нейрона игнорирует многие свойства своего биологического двойника. Например, она не принимает во внимание задержки во времени, которые воздействуют на динамику системы. Входные сигналы сразу же порождают выходной сигнал. И, что более важно, она не учитывает воздействий функции частотной модуляции или синхронизирующей функции биологического нейрона, которые ряд исследователей считают решающими.
Несмотря на эти ограничения, сети, построенные из этих нейронов, обнаруживают свойства, сильно напоминающие биологическую систему. Только время и исследования смогут ответить на вопрос, являются ли подобные совпадения случайными или следствием того, что в модели верно схвачены важнейшие черты биологического нейрона.